![線粒體[線粒體]](/img/e/3a9/nBnauM3XzETNzkTM5czNzQzN1UTM1QDN5MjM5ADMwAjMwUzL3czLzUzLt92YucmbvRWdo5Cd0FmL0E2LvoDc0RHa.jpg "線粒體[線粒體]")
形態特徵
大小
 線粒體結構示意圖
線粒體結構示意圖 線粒體是一些大小不一的球狀、棒狀或細絲狀顆粒,一般為0.5-1.0μm,長1-2μm,在光學顯微鏡下,需用特殊的染色,才能加以辨別。 在動物細胞中,線粒體大小受細胞代謝水平限制。不同組織在不同條件下可能產生體積異常膨大的線粒體,稱為“巨線粒體”(megamitochondria):胰臟外分泌細胞中可長達10-20μm;神經元胞體中的線粒體尺寸差異很大,有的也可能長達10μm;人類成纖維細胞的線粒體則更長,可達40μm。有研究表明在低氧氣分壓的環境中,某些如菸草的植物的線粒體能可逆地變為巨線粒體,長度可達80μm,並形成網路。
形狀
 圖中編號為9的是線粒體
圖中編號為9的是線粒體 線粒體一般呈短棒狀或圓球狀,但因生物種類和生理狀態而異,還可呈環狀、線狀、啞鈴狀、分杈狀、扁盤狀或其它形狀。成型蛋白(shape-forming protein)介導線粒體以不同方式與周圍的細胞骨架接觸或線上粒體的兩層膜間形成不同的連線可能是線粒體在不同細胞中呈現出不同形態的原因。
數量
不同生物的不同組織中線粒體數量的差異是巨大的。有許多細胞擁有多達數千個的線粒體(如肝臟細胞中有1000-2000個線粒體),而一些細胞則只有一個線粒體(如酵母菌細胞的大型分支線粒體)。大多數哺乳動物的成熟紅細胞不具有線粒體。一般來說,細胞中線粒體數量取決於該細胞的代謝水平,代謝活動越旺盛的細胞線粒體越多。
分布
線粒體分布方向與微管一致,通常分布在細胞功能旺盛的區域:如在腎臟細胞中靠近微血管,呈平行或柵狀排列;在腸表皮細胞中呈兩極分布,集中在頂端和基部;在精子中分布在鞭毛中區。在卵母細胞體外培養中,隨著細胞逐漸成熟,線粒體會由在細胞周邊分布發展成均勻分布。線粒體在細胞質中能以微管為導軌、由馬達蛋白提供動力向功能旺盛的區域遷移。
組成
![線粒體[線粒體]](/img/5/7d7/nBnauM3X1MjMzUTN0IDOzQzN1UTM1QDN5MjM5ADMwAjMwUzLygzL3IzLt92YucmbvRWdo5Cd0FmLwE2LvoDc0RHa.jpg) 線粒體[線粒體]
線粒體[線粒體] 線粒體的化學組分主要包括水、蛋白質和脂質,此外還含有少量的輔酶等小分子及核酸。蛋白質占線粒體乾重的65-70%。線粒體中的蛋白質既有可溶的也有不溶的。可溶的蛋白質主要是位於線粒體基質的酶和膜的外周蛋白;不溶的蛋白質構成膜的本體,其中一部分是鑲嵌蛋白,也有一些是酶。線粒體中脂類主要分布在兩層膜中,占乾重的20-30%。線上粒體中的磷脂占總脂質的3/4以上。同種生物不同組織線粒體膜中磷脂的量相對穩定。含豐富的心磷脂和較少的膽固醇是線粒體在組成上與細胞其他膜結構的明顯差別。
結構
線粒體由外至內可劃分為線粒體外膜(OMM)、線粒體膜間隙、線粒體內膜(IMM)和線粒體基質四個功能區。處於線粒體外側的膜彼此平行,都是典型的單位膜。其中,線粒體外膜較光滑,起細胞器界膜的作用;線粒體內膜則向內皺褶形成線粒體嵴,負擔更多的生化反應。這兩層膜將線粒體分出兩個區室,位於兩層線粒體膜之間的是線粒體膜間隙,被線粒體內膜包裹的是線粒體基質。
研究歷史
![線粒體[線粒體]](/img/e/1ad/nBnauM3XyQzMwcjM0YTOzQzN1UTM1QDN5MjM5ADMwAjMwUzL2kzL0IzLt92YucmbvRWdo5Cd0FmLzE2LvoDc0RHa.jpg) 線粒體[線粒體]
線粒體[線粒體] 線粒體的研究是從19世紀50年代末開始的。
1857年,瑞士解剖學家及生理學家阿爾伯特·馮·科立克在肌肉細胞中發現了顆粒狀結構。另外的一些科學家在其他細胞中也發現了同樣的結構,證實了科立克的發現。德國病理學家及組織學家理察·阿爾特曼將這些顆粒命名為“原生粒”(bioblast)並於1886年發明了一種鑑別這些顆粒的染色法。阿爾特曼猜測這些顆粒可能是共生於細胞內的獨立生活的細菌。
1898年,德國科學家卡爾·本達因這些結構時而呈線狀時而呈顆粒狀,所以用希臘語中“線”和“顆粒”對應的兩個詞——“ mitos”和“ chondros”——組成“ mitochondrion”來為這種結構命名,這個名稱被沿用至今。一年後,美國化學家萊昂諾爾·米歇利斯開發出用具有還原性的健那綠染液為線粒體染色的方法,並推斷線粒體參與某些氧化反應。這一方法於1900年公布,並由美國細胞學家埃德蒙·文森特·考德里推廣。德國生物化學家奧托·海因里希·沃伯格成功完成線粒體的粗提取且分離得到一些催化與氧有關的反應的呼吸酶,並提出這些酶能被氰化物(如氫氰酸)抑制的猜想。
英國生物學家大衛·基林在1923年至1933年這十年間對線粒體內的氧化還原鏈(redox chain)的物質基礎進行探索,辨別出反應中的電子載體——細胞色素。
沃伯格於1931年因“發現呼吸酶的性質及作用方式”被授予諾貝爾生理學或醫學獎。
美國維吉尼亞大學最新一項研究表明,動植物細胞中的線粒體其實是寄生細菌,早期寄生細菌可以對動物和植物提供能量,在細胞中作為能量寄生蟲存在,對寄居體十分有益。新一代DNA序列技術解碼18種細菌基因組,這些細菌是線粒體的近親生物。
主要結構
外膜
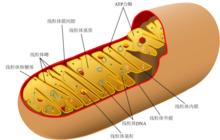 線粒體剖面圖
線粒體剖面圖 線粒體外膜是位於線粒體最外圍的一層單位膜,厚度約為6-7nm。其中磷脂與蛋白質的質量為0.9:1,與真核細胞細胞膜的同一比例相近。線粒體外膜中酶的含量相對較少,其標誌酶為單胺氧化酶。線粒體外膜包含稱為“孔蛋白”的整合蛋白,其內部通道寬約2-3nm,這使線粒體外膜對分子量小於5000Da的分子完全通透。分子量大於上述限制的分子則需擁有一段特定的信號序列以供識別並通過外膜轉運酶(translocase of the outer membrane,TOM)的主動運輸來進出線粒體。線粒體外膜主要參與諸如脂肪酸鏈延伸、腎上腺素氧化以及色氨酸生物降解等生化反應,它也能同時對那些將線上粒體基質中進行徹底氧化的物質先行初步分解。細胞凋亡過程中,線粒體外膜對多種存在於線粒體膜間隙中的蛋白的通透性增加,使致死性蛋白進入細胞質基質,促進了細胞凋亡。高分辨三維X射線攝影可見內質網及線粒體之間的有20%膜是緊密接觸的,在這些接觸位點上線粒體外膜與內質網膜通過某些蛋白質相連,形成稱為“線粒體結合內質網膜”(mitochondria-associated ER-membrane,MAM)的結構。該結構在脂質的相互交換和線粒體與內質網間的鈣離子信號傳導等過程中都有重要作用。
膜間隙
線粒體膜間隙是線粒體外膜與線粒體內膜之間的空隙,寬約6-8nm,其中充滿無定形液體。由於線粒體外膜含有孔蛋白,通透性較高,而線粒體內膜通透性較低,所以線粒體膜間隙內容物的組成與細胞質基質十分接近,含有眾多生化反應底物、可溶性的酶和輔助因子等。線粒體膜間隙中還含有比細胞質基質中濃度更高的腺苷酸激酶、單磷酸激酶和二磷酸激酶等激酶,其中腺苷酸激酶是線粒體膜間隙的標誌酶。線粒體膜間隙中存在的蛋白質可統稱為“線粒體膜間隙蛋白質”,這些蛋白質全部在細胞質基質中合成。
內膜
線粒體內膜是位於線粒體外膜內側、包裹著線粒體基質的單位膜。線粒體內膜中蛋白質與磷脂的質量比約為0.7 :0.3,並含有大量的心磷脂(心磷脂常為細菌細胞膜的成分)。線粒體內膜的某些部分會向線粒體基質摺疊形成線粒體嵴。線粒體內膜的標誌酶是細胞色素氧化酶。
線粒體通過向內凹形成嵴,從而來增加內膜面積。然後是更多的反應能在內膜上進行。
線粒體內膜含有比外膜更多的蛋白質(超過151種,約占線粒體所含所有蛋白質的五分之一),所以承擔著更複雜的生化反應。存在於線粒體內膜中的幾類蛋白質主要負責以下生理過程:特異性載體運輸磷酸、谷氨酸、鳥氨酸、各種離子及核苷酸等代謝產物和中間產物;內膜轉運酶(translocase of the inner membrane,TIM)運輸蛋白質;參與氧化磷酸化中的氧化還原反應;參與ATP的合成;控制線粒體的分裂與融合。
嵴
線粒體嵴簡稱“嵴”,是線粒體內膜向線粒體基質折褶形成的一種結構。線粒體嵴的形成增大了線粒體內膜的表面積。在不同種類的細胞中,線粒體嵴的數目、形態和排列方式可能有較大差別。線粒體嵴主要有幾種排列方式,分別稱為“片狀嵴”(lamellar cristae)、“管狀嵴”(tubular cristae)和“泡狀嵴”(vesicular cristae)。片狀排列的線粒體嵴主要出現在高等動物細胞的線粒體中,這些片狀嵴多數垂直於線粒體長軸;管狀排列的線粒體嵴則主要出現在原生動物和植物細胞的線粒體中。有研究發現,睪丸間質細胞中既存在層狀嵴也存在管狀嵴。線粒體嵴上有許多有柄小球體,即線粒體基粒,基粒中含有ATP合酶,能利用呼吸鏈產生的能量合成三磷酸腺苷。所以需要較多能量的細胞,線粒體嵴的數目一般也較多。但某些形態特殊的線粒體嵴由於沒有ATP合酶,所以不能合成ATP。
基質
 膜結構
膜結構 線粒體基質是線粒體中由線粒體內膜包裹的內部空間,其中含有參與三羧酸循環、脂肪酸氧化、胺基酸降解等生化反應的酶等眾多蛋白質,所以較細胞質基質黏稠。蘋果酸脫氫酶是線粒體基質的標誌酶。線粒體基質中一般還含有線粒體自身的DNA(即線粒體DNA)、RNA和核糖體(即線粒體核糖體)。
線粒體DNA是線粒體中的遺傳物質,呈雙鏈環狀。一個線粒體中可有一個或數個線粒體DNA分子。線粒體RNA是線粒體DNA的表達產物,RNA編輯也普遍存在於線粒體RNA中,是線粒體產生功能蛋白所必不可少的過程。線粒體核糖體是存在於線粒體基質內的一種核糖體,負責完成線粒體內進行的翻譯工作。線粒體核糖體的沉降係數介乾55S-56S之間。一般的線粒體核糖體由28S核糖體亞基(小亞基)和39S核糖體亞基(大亞基)組成。在這類核糖體中,rRNA約占25%,核糖體蛋白質約占75%。線粒體核糖體是已發現的蛋白質含量最高的一類核糖體。線粒體基質中存在的蛋白質統稱為“線粒體基質蛋白質”,包括DNA聚合酶、RNA聚合酶、檸檬酸合成酶以及三羧酸循環酶系中的酶類。大部分線粒體基質蛋白是由核基因編碼的。線粒體基質蛋白不一定只線上粒體基質中表達,它們也可以線上粒體外表達。
主要功能
能量轉化
線粒體是真核生物進行氧化代謝的部位,是糖類、脂肪和胺基酸最終氧化釋放能量的場所。線粒體負責的最終氧化的共同途徑是三羧酸循環與氧化磷酸化,分別對應有氧呼吸的第二、三階段。細胞質基質中完成的糖酵解和線上粒體基質中完成的三羧酸循環在會產還原型煙醯胺腺嘌呤二核苷酸(reduced nicotinarnide adenine dinucleotide,NADH)和還原型黃素腺嘌呤二核苷酸(reduced flavin adenosine dinucleotide,FADH2)等高能分子,而氧化磷酸化這一步驟的作用則是利用這些物質還原氧氣釋放能量合成ATP。在有氧呼吸過程中,1分子葡萄糖經過糖酵解、三羧酸循環和氧化磷酸化將能量釋放後,可產生30-32分子ATP(考慮到將NADH運入線粒體可能需消耗2分子ATP)。如果細胞所在環境缺氧,則會轉而進行無氧呼吸。此時,糖酵解產生的丙酮酸便不再進入線粒體內的三羧酸循環,而是繼續在細胞質基質中反應(被NADH還原成乙醇或乳酸等發酵產物),但不產生ATP。所以在無氧呼吸過程中,1分子葡萄糖只能在第一階段產生2分子ATP。
三羧酸循環
糖酵解中生成的每分子丙酮酸會被主動運輸轉運穿過線粒體膜。進入線粒體基質後,丙酮酸會被氧化,並與輔酶A結合生成CO2、還原型輔酶Ⅰ和乙醯輔酶A。乙醯輔酶A是三羧酸循環(也稱為“檸檬酸循環”或“Krebs循環”)的初級底物。參與該循環的酶除位於線粒體內膜的琥珀酸脫氫酶外都游離於線粒體基質中。在三羧酸循環中,每分子乙醯輔酶A被氧化的同時會產生起始電子傳遞鏈的還原型輔因子(包括3分子NADH和1分子FADH2)以及1分子三磷酸鳥苷(GTP)。
氧化磷酸化
![線粒體[線粒體]](/img/e/e9e/nBnauM3XzEjMwMTM2QDOzQzN1UTM1QDN5MjM5ADMwAjMwUzL0gzLyQzLt92YucmbvRWdo5Cd0FmLyE2LvoDc0RHa.jpg) 線粒體[線粒體]
線粒體[線粒體] NADH和FADH2等具有還原性的分子(在細胞質基質中的還原當量可從由逆向轉運蛋白構成的蘋果酸-天冬氨酸穿梭系統或通過磷酸甘油穿梭作用進入電子傳遞鏈)在電子傳遞鏈裡面經過幾步反應最終將氧氣還原並釋放能量,其中一部分能量用於生成ATP,其餘則作為熱能散失。線上粒體內膜上的酶複合物(NADH-泛醌還原酶、泛醌-細胞色素c還原酶、細胞色素c氧化酶)利用過程中釋放的能量將質子逆濃度梯度泵入線粒體膜間隙。雖然這一過程是高效的,但仍有少量電子會過早地還原氧氣,形成超氧化物等活性氧(ROS),這些物質能引起氧化應激反應使線粒體性能發生衰退。
當質子被泵入線粒體膜間隙後,線粒體內膜兩側便建立起了電化學梯度,質子就會有順濃度梯度擴散的趨勢。質子唯一的擴散通道是ATP合酶(呼吸鏈複合物V)。當質子通過複合物從膜間隙回到線粒體基質時,電勢能被ATP合酶用於將ADP和磷酸合成ATP。這個過程被稱為“化學滲透”,是一種協助擴散。彼得·米切爾就因為提出了這一假說而獲得了1978年諾貝爾獎。1997年諾貝爾獎獲得者保羅·博耶和約翰·瓦克闡明了ATP合酶的機制。
儲存鈣離子
線粒體可以儲存鈣離子,可以和內質網、細胞外基質等結構協同作用,從而控制細胞中的鈣離子濃度的動態平衡。線粒體迅速吸收鈣離子的能力使其成為細胞中鈣離子的緩衝區。線上粒體內膜膜電位的驅動下,鈣離子可由存在於線粒體內膜中的單向運送體輸送進入線粒體基質;排出線粒體基質時則需要鈉-鈣交換蛋白的輔助或通過鈣誘導鈣釋放(calcium-induced-calcium-release,CICR)機制。在鈣離子釋放時會引起伴隨著較大膜電位變化的“鈣波”(calcium wave),能激活某些第二信使系統蛋白,協調諸如突觸中神經遞質的釋放及內分泌細胞中激素的分泌。線粒體也參與細胞凋亡時的鈣離子信號轉導。
其他功能
除了合成ATP為細胞提供能量等主要功能外,線粒體還承擔了許多其他生理功能。
 顯微鏡下
顯微鏡下 ·調節膜電位並控制細胞程式性死亡:當線粒體內膜與外膜接觸位點處生成了由己糖激酶(細胞質基質蛋白)、外周苯並二氮受體和電壓依賴陰離子通道(線粒體外膜蛋白)、肌酸激酶(線粒體膜間隙蛋白)、ADP-ATP載體(線粒體內膜蛋白)和親環蛋白D(線粒體基質蛋白)等多種蛋白質組成的通透性轉變孔道(PT孔道)後,會使線粒體內膜通透性提高,引起線粒體跨膜電位的耗散,從而導致細胞凋亡。線粒體膜通透性增加也能使誘導凋亡因子(AIF)等分子釋放進入細胞質基質,破壞細胞結構。
·細胞增殖與細胞代謝的調控;
·合成膽固醇及某些血紅素。
線粒體的某些功能只有在特定的組織細胞中才能展現。例如,只有肝臟細胞中的線粒體才具有對氨氣(蛋白質代謝過程中產生的廢物)造成的毒害解毒的功能。
起源學說
對於線粒體的起源有兩種假說,分別為內共生學說與非內共生學說:
內共生學說
該學說認為線粒體起源於被另一個細胞吞噬的線粒體祖先——原線粒體——一種能進行三羧酸循環和電子傳遞的革蘭氏陰性菌。這種好氧細菌是變形菌門下的一個分支,與立克次氏體有密切關係。原線粒體被吞噬後,並沒有被消化,而是與宿主細胞形成了共生關係——寄主可以從宿主處獲得更多營養,而宿主則可使用寄主產生的能量——這種關係增加了細胞的競爭力,使其可以適應更多的生存環境。在長期對寄主和宿主都有利的互利共生中,原線粒體逐漸演變形成了線粒體 ,使宿主細胞中進行的糖酵解和原線粒體中進行的三羧酸循環和氧化磷酸化成功耦合。有研究認為,這種共生關係大約發生在17億年以前,與進化趨異產生真核生物和古細菌的時期幾乎相同。但線粒體與真核生物細胞核出現的先後關係仍存在爭議。
現已發現支持內共生學說的證據包括 :
遺傳信息轉移:近期的分子生物學和生物信息學的研究發現真核細胞的細胞核中存在可能屬於呼吸細菌或藍細菌的遺傳信息,說明最初的呼吸細菌和藍細菌的大部分基因組在漫長的共進化過程中發生了向細胞核的轉移。
線粒體基因組與細菌基因組具有明顯的相似性:包括1)線粒體擁有自己DNA,其形狀與細菌的環狀DNA類似,無組蛋白結合;2)鹼基比例、核苷酸序列、基因結構特徵等方面相似,不含5mC;3)線粒體具有自身的DNA聚合酶及RNA聚合酶,能進行獨立的複製與轉錄;4)其mRNA、rRNA的沉降係數與細菌的相似。
線粒體具備獨立、完整的蛋白質合成系統:與真核細胞的蛋白質合成系統相比,線粒體蛋白質合成的多數特徵與細菌蛋白質合成系統更相似,包括1)蛋白質合成從N-甲醯甲硫氨酸開始,而真核細胞從甲硫氨酸開始;2)線粒體的核糖體小於真核生物的80S核糖體;3)線粒體、葉綠體、原核生物中存在5SrRNA,而不少真核生物的核糖體中存在5.8SrRNA;4)線粒體中的蛋白質合成因子具有原核生物核糖體的識別特異性,但不能識別細胞質核糖體;5)線粒體mRNA與線粒體核糖體形成多核糖體;6)線粒體、葉綠體上的蛋白質合成可被氯黴素、四環素所抑制,而抑制真核生物蛋白質合成的放線菌酮則對他們無抑制作用;7)線粒體的RNA聚合酶可被原核細胞RNA聚合酶抑制劑利福黴素所抑制,但不被真核細胞RNA聚合酶抑制劑放線菌素D所抑制等。
線粒體分裂方式與細菌相似:線粒體及葉綠體均以縊裂的方式分裂增殖,類似於細菌。
線粒體的膜特性:線粒體外膜與真核細胞內膜相似,線粒體內膜與細菌質膜相似;線粒體內膜的蛋白質/脂質的比例遠大於外膜,與細菌相似。
其他特徵:線粒體的磷脂成分、呼吸類型和Cyt c的初級結構均與反硝化副球菌或紫色非硫光合細菌非常接近,暗示線粒體的祖先可能是這兩種菌的一種。
遺傳密碼比較:線粒體的遺傳密碼與變形菌門細菌的遺傳密碼更為相似;
1.遺傳信息轉移
2.線粒體基因組與細菌基因組具有明顯的相似性
3.線粒體具備獨立、完整的蛋白質合成系統:
4.線粒體分裂方式與細菌相似
5.線粒體的膜特性
6.其他特徵
7.遺傳密碼比較
不足之處:
從進化角度,如何解釋在代謝上明顯占優勢的共生體反而將大量的遺傳信息轉移到宿主細胞中?
不能解釋細胞核是如何進化來的,即原核細胞如何演化為真核細胞?
線粒體和葉綠體的基因組中存在內含子,而真細菌原核生物基因組中不存在內含子,如果同意內共生起源學說的觀點,那么線粒體和葉綠體基因組中的內含子從何發生?
非內共生學說
非內共生學說又稱為“細胞分化學說”,認為線粒體的發生是由細胞膜或內質網膜等生物膜系統中的膜結構演變而來的。非內共生學說有幾種模型,主流的模型認為在細胞進化的最初階段,原核細胞基因組複製後並不伴有典型的無絲分裂,而是擬核附近的細胞膜內陷形成雙層膜,將其中一個基因組包圍、隔離,進而發生細胞分裂。未分裂出來的子細胞則緩慢演化為細胞核、線粒體和葉綠體等高度特化的細胞結構。
不足之處:
實驗證據不多
無法解釋為何線粒體、葉綠體與細菌在DNA分子結構和蛋白質合成性能上有那么多相似之處
對線粒體和葉綠體的DNA酶、RNA酶和核糖體的來源也很難解釋。
真核細胞的細胞核能否起源於細菌的核區?
遺傳學
線粒體基因組
線粒體基因組中基因的數量很少,規模遠小於細菌基因組。但內共生學說認為線粒體源於被吞噬的細菌,那么兩者基因組規模應該較為相似。為了解釋這一現象,有猜想認為原線粒體的基因除了丟失了一些外,大部分轉移到了宿主細胞的細胞核中,所以核基因編碼了在超過98%的線粒體表達內的蛋白質。某些有線粒體,但線粒體中不含DNA的生物(如隱孢子蟲等)的mtDNA可能已完全丟失或整合入核DNA中。線粒體DNA(mtDNA)線上粒體中有2-10個備份,呈雙鏈環狀(但也有呈線狀的特例存在)。mtDNA長度一般為幾萬至數十萬鹼基對,人類mtDNA的長度為16,569bp,擁有有37個基因,編碼了兩種rRNA(12S rRNA和16S rRNA)、22種tRNA(同樣轉運20種標準胺基酸,只是亮氨酸和絲氨酸都有兩種對應的tRNA)以及13種多肽(呼吸鏈複合物Ⅰ、Ⅲ、Ⅳ、Ⅴ的亞基)。mtDNA的長度和線粒體基因組的大小因物種而異,表一列出了幾種模式生物mtDNA的長度:
| 生物 | 學名 | mtDNA長度(bp) |
| 芽殖酵母 | Saccharomyces cerevisiae | 85779 |
| 裂殖酵母 | Schizosaccharomyces pombe | 19431 |
| 擬南芥 | Arabidopsis thaliana | 366924 |
| 水稻 | Oryza sativa | 490520 |
| 秀麗隱桿線蟲 | Caenorhabditis elegans | 13794 |
| 黑腹果蠅 | Drosophila melanogaster | 19517 |
| 非洲爪蟾 | Xenopus laevis | 17553 |
| 小鼠 | Mus musculus | 16300 |
mtDNA利用率極高,線粒體基因組各基因之間排列十分緊湊,部分區域還可能出現重疊(即前一個基因的最後一段鹼基與下一個基因的第一段鹼基相銜接)。人類mtDNA中基因間隔區總共只有87bp,占mtDNA總長的0.5%。mtDNA的兩條DNA單鏈均有編碼功能,其中重鏈編碼兩個rRNA、12個mRNA和14個tRNA;輕鏈編碼一個mRNA和8個tRNA。mtDNA一般沒有內含子(如人類的mtDNA等),但也已發現某些真核生物的mtDNA擁有內含子,這些生物包括:盤基網柄菌等原生生物和酵母菌(其OXi3基因有9個內含子)。這些mtDNA中的內含子在基因轉錄產物的加工和翻譯中可能有一定功能。
線粒體基因組通常都是存在於同一個mtDNA分子中,但少數生物的線粒體基因組卻分別儲存在多個不同的mtDNA中。例如,人虱的線粒體基因組就分開儲藏於18個長約3-4kb的微型環狀DNA中,每個DNA分子只分配到了1-3個基因。這些微型環狀DNA之間也存在著同源或非同源的基因重組現象,但成因未知。
2019年3月,發表在PNAS 《美國科學院院刊》上的研究表明,線粒體可由父系遺傳。來自美國辛辛那提兒童醫院的黃濤生博士和梅奧診所的Paldeep Atwal博士稱他們在三個家庭中發現了mtDNA雙親遺傳。傳統觀念里,大多數哺乳動物的線粒體和線粒體DNA都是只通過母系遺傳。儘管有其他物種已被發現線粒體偶爾會經歷父系遺傳,但之前關於人類父系遺傳線粒體的報導大多是因為污染或樣本混淆。然而,2019年美國的實驗室發表論文,稱他們在三個家庭中發現了mtDNA雙親遺傳。研究人員還在獨立實驗室中通過不同方法證實了他們的成果 。
遺傳密碼
線粒體中擁有一套獨特的遺傳系統。在進行人類線粒體遺傳學研究時,人們確認線粒體的遺傳密碼與通用遺傳密碼也有些許差異。自從上述發現證明並不只存在單獨的一種遺傳密碼之後,許多有輕微不同的遺傳密碼都陸續連發現。線上粒體的遺傳密碼中最常見的差異是:AUA由終止密碼子變為甲硫氨酸的密碼子、UGA由終止密碼子變為色氨酸的密碼子、AGA和AGG由精氨酸的密碼子變為終止密碼子(植物等生物的線粒體遺傳密碼另有差異,參見表二)。此外,也有某些特例是只涉及終止密碼子的,在山羊支原體線粒體遺傳密碼的UGA由終止密碼子變為色氨酸的密碼子,而且使用頻率比UGG更高;四膜蟲線粒體遺傳密碼里只有UGA一種終止密碼子,其UAA和UAG由終止密碼子變為谷氨醯胺的密碼子;而游仆蟲線粒體遺傳密碼里則只有UAA和UAG兩種終止密碼子,其UGA由終止密碼子變為半胱氨酸的密碼子。通過線粒體遺傳密碼和通用遺傳密碼的對比,可以推導出遺傳密碼演化過程的可能模式。
| 密碼子 | 通用密碼 | 線粒體遺傳密碼 | |||
| 真菌 | 植物 | 無脊椎動物 | 哺乳動物 | ||
| UGA | 終止密碼子 | 色氨酸 | 終止密碼子 | 色氨酸 | 色氨酸 |
| AUA | 異亮氨酸 | 甲硫氨酸 | 異亮氨酸 | 甲硫氨酸 | 甲硫氨酸 |
| CUA | 亮氨酸 | 蘇氨酸 | 亮氨酸 | 亮氨酸 | 亮氨酸 |
| AGA、AGG | 精氨酸 | 精氨酸 | 精氨酸 | 絲氨酸 | 終止密碼子 |
分裂與融合
線粒體的融合是與分裂協同進行的,過程高度保守,需要在多種蛋白質的精確調控下完成。兩者一般保持動態平衡,這種平衡對維持線粒體正常的形態、分布和功能十分重要。線粒體融合與分裂間的失衡可產生巨型線粒體,這種過大的線粒體常見於病變的肝細胞、惡性營養不良患者的胰臟細胞和白血病患者骨髓的巨噬細胞中。分裂異常會導致線粒體破碎,而融合異常則會導致線粒體形態延長,兩者都會影響線粒體的功能。分裂與融合活動異常的線粒體膜電位通常會降低,並最終經線粒體自噬作用清除。
線粒體的分裂在真核細胞內經常發生。為了保證在細胞發生分裂後每個子細胞都能繼承母細胞的線粒體,母細胞中的線粒體在一個細胞周期需要至少複製一次。即使是在不再分裂的細胞內,線粒體為了填補已老化的線粒體造成的空缺也需要進行分裂。的線粒體以與細菌的無絲分裂類似的方式進行增殖,可細分為三種模式:
間壁分離(見於部分動物和植物線粒體):線粒體內部首先由內膜形成隔,隨後外膜的一部分內陷,插入到隔的雙層膜之間,將線粒體一分為二。
收縮分離(見於蕨類植物和酵母菌線粒體):線粒體中部先縊縮同時向兩端不斷拉長然後一分為二。
出芽分離(見於蘚類植物和酵母菌線粒體):線粒體上先出現小芽,小芽脫落後成長、發育為成熟線粒體。
線粒體的融合也是細胞中的基本事件,對線粒體正常功能的發揮具有非常重要的作用。人類細胞需要通過線粒體融合的互補作用來抵抗衰老;酵母細胞線粒體融合發生障礙會引起呼吸鏈缺陷。線粒體間的融合需在一種分子量約為800kDa的蛋白質複合物——“融合裝置”(fusion machinery)的介導下進行,該過程可大致分為四個步驟:錨定、外膜融合、內膜融合以及基質內含物融合。
群體遺傳學
因為mtDNA幾乎不發生基因重組,所以遺傳學家長期將其作為研究群體遺傳學與進化生物學的信息來源。所有mtDNA是以單一單元(單體型)進行遺傳的(而不像細胞核中的DNA儲存在多個染色體中),它們在親本與子代之間的傳遞關係並不複雜,因此不同個體間mtDNA的聯繫便可以利用系統發生樹來表現。而從這些系統發生樹的形態中人們可以得知種群的進化史。人類進化遺傳學中運用分子鐘技術推算出了線粒體夏娃最晚出現的時間(這個成果被認為是人類由非洲單地起源的有力依據)是利用mtDNA研究群體遺傳學的典型例子。另外一個例子是對尼安德特人骨骼化石中mtDNA測序。該測序的結果顯示,尼安德特人與解剖學意義上的現代人在mtDNA序列上有較大差異,說明兩者間缺乏基因交流。雖然mtDNA在遺傳學研究中占據了重要地位,但是mtDNA序列中的信息只能反映所考察的群體中的雌性成員的演化進程,而不能代表整個種群。這一缺陷需要由對父系遺傳序列(如Y染色體上的非重組區)的測序彌補。廣義上來說,只有既考慮了mtDNA又考慮了核DNA的遺傳學研究才能為種群的進化史提供全面的線索。
病理
線粒體是對各種損傷最為敏感的細胞器之一。在細胞損傷時最常見的病理改變可概括為線粒體數量、大小和結構的改變:
數量的改變
線粒體的平均壽命約為10天。衰亡的線粒體可通過保留的線粒體直接分裂為二予以補充。在病理狀態下,線粒體的增生實際上是對慢性非特異性細胞損傷的適應性反應或細胞功能升高的表現。例如心瓣膜病時的心肌線粒體、周圍血液循環障礙伴間歇性跛行時的骨骼肌線粒體的呈增生現象。
線粒體數量減少則見於急性細胞損傷時線粒體崩解或自溶的情況下,持續約15分鐘。慢性損傷時由於線粒體逐漸增生,故一般不見線粒體減少(甚至反而增多)。此外,線粒體的減少也是細胞未成熟和(或)去分化的表現。
大小改變
細胞損傷時最常見的改變為線粒體腫大。根據線粒體的受累部位可分為基質型腫脹和嵴型腫脹二種類型,而以前者為常見。基質型腫脹時線粒體變大變圓,基質變淺、嵴變短變少甚至消失(圖1-9)。在極度腫脹時,線粒體可轉化為小空泡狀結構。此型腫脹為細胞水腫的部分改變。光學顯微鏡下所謂的濁腫細胞中所見的細顆粒即腫大的線粒體。嵴型腫較少見,此時的腫脹局限於嵴內隙,使扁平的嵴變成燒瓶狀乃至空泡狀,而基質則更顯得緻密。嵴型腫脹一般為可復性,但當膜的損傷加重時,可經過混合型而過渡為基質型。
線粒體為對損傷極為敏感的細胞器,其腫脹可由多種損傷因子引起,其中最常見的為缺氧;此外,微生物毒素、各種毒物、射線以及滲透壓改變等亦可引起。但輕度腫大有時可能為其功能升高的表現,較明顯的腫脹則恆為細胞受損的表現。但只要損傷不過重、損傷因子的作用不過長,腫脹仍可恢復。
線粒體的增大有時是器官功能負荷增加引起的適應性肥大,此時線粒體的數量也常增多,例如見於器官肥大時。反之,器官萎縮時,線粒體則縮小、變少。
結構的改變
線粒體嵴是能量代謝的明顯指征,但嵴的增多未必均伴有呼吸鏈酶的增加。嵴的膜和酶平行增多反映細胞的功能負荷加重,為一種適應狀態的表現;反之,如嵴的膜和酶的增多不相平行,則是胞漿適應功能障礙的表現,此時細胞功能並不升高。
在急性細胞損傷時(大多為中毒或缺氧),線粒體的嵴被破壞;慢性亞致死性細胞損傷或營養缺乏時,線粒體的蛋白合成受障,以致線粒體幾乎不再能形成新的嵴。
根據細胞損傷的種類和性質,可線上粒體基質或嵴內形成病理性包含物。這些包含物有的呈晶形或副晶形(可能由蛋白構成),如線上粒體性肌病或進行性肌營養不良時所見,有的呈無定形的電子緻密物,常見於細胞趨於壞死時,乃線粒體成分崩解的產物(脂質和蛋白質),被視為線粒體不可復性損傷的表現。線粒體損傷的另一種常見改變為髓鞘樣層狀結構的形成,這是線粒體膜損傷的結果。
衰亡或受損的線粒體,最終由細胞的自噬過程加以處理並最後被溶酶體酶所降解消化。
線粒體與衰老
線粒體是直接利用氧氣製造能量的部位,90%以上吸入體內的氧氣被線粒體消耗掉。但是,氧是個“雙刃劍”,一方面生物體利用氧分子製造能量,另一方面氧分子在被利用的過程中會產生極活潑的中間體(活性氧自由基)傷害生物體造成氧毒性。生物體就是在不斷地與氧毒性進行鬥爭中求得生存和發展的,氧毒性的存在是生物體衰老的最原初的原因。線粒體利用氧分子的同時也不斷受到氧毒性的傷害,線粒體損傷超過一定限度,細胞就會衰老死亡。生物體總是不斷有新的細胞取代衰老的細胞以維持生命的延續,這就是細胞的新陳代謝。
線粒體與疾病
人類線粒體出現問題會導致線粒體病,線粒體病是一大類遺傳代謝病,線粒體病主要包括:母系遺傳Leigh綜合徵、線粒體肌病、多系統疾病、心肌病、進行性眼外肌麻痹、Leer遺傳性視神經病、線粒體肌病,肌病,糖尿病和耳聾、共濟失調舞蹈病、細胞外基質慢性遊走性紅斑、進行性眼外肌麻痹、肌紅蛋白尿電機神經元疾病,鐵粒幼細胞貧血、MERRF-線粒體肌病、肌陣攣(癲癇)、線粒體腦肌病、MERRF、線粒體肌病、共濟失調並發色素性視網膜炎、家族性雙側紋狀體壞死、共濟失調並發色素性視網膜炎、家族性雙側紋狀體壞死、骨骼肌溶解症、嬰兒猝死綜合徵等等疾病。
線粒體病遺傳方式複雜,導致疾病的原因主要由核基因和線粒體基因造成,臨床表現複雜,確切病因的診斷十分困難,往往通過大分子酶學活性檢測分析並結合遺傳學基因分析的雙重手段確定病因。
線粒體基因組屬於母系遺傳,為了避免新生兒缺陷,產前媽媽的線粒體基因組分析十分必要。
2014年7月,科學家發現促進癌症轉移的線粒體開關,線粒體是細胞的能量工廠,當腫瘤細胞中線粒體的功能發生改變時就會促進細胞的遷移,最終導致腫瘤成功轉移。研究人員測定了腫瘤細胞中線粒體促進腫瘤轉移過程中涉及的分子機制,結果發現,在特定的條件下,線粒體可以產生過多的超氧離子自由基,超氧離子的過量產生就會引發腫瘤轉移灶的形成,最終腫瘤轉移組織就會在新的組織中形成腫瘤。
線粒體實驗
染色觀察
線粒體——示教:3號片
小狗胰臟,Regaud氏液固定,石蠟切片,鐵蘇木素染色。
線粒體用鐵蘇木素染色呈黑色,分布於核周圍的細胞質中,線粒體在高倍鏡下呈粒狀、線狀或短棒狀,或直或曲,輪廓鮮明。
胰臟的分泌細胞呈錐形,核大而圓,位於細胞中央,細胞游離端聚集有許多大而圓的黑色顆粒為分泌顆粒。
提取觀察
線粒體是細胞中重要的細胞器,存在於絕大多數生活細胞中,它的主要功能是提供細胞內各種物質代謝所需要的能量。正由於這樣,對線粒體膜,呼吸鏈酶及線粒體DNA等成分的結構,功能以及物理化學性質的研究已經成為細胞生物學研究中的重要課題,所以提取線粒體的技術已經成為線粒體研究中必不可少的手段,線粒體大量存在於代謝旺盛的細胞中,如動物的心肌,肝,腎等器官和組織的細胞中,大量置備線粒體就是從這些器官組織中提取,當所用樣品較少時(如電鏡和光鏡的觀察)可採用從組織培養細胞中提取,本實驗就是介紹兩種材料製備用於光鏡觀察的線粒體。
一、目的與要求
了解提取線粒體的基本原理及其過程,通過光學顯微鏡的觀察了解體外分離的線粒體的一般形態。
二、基本原理
線粒體具有完整的結構,一定的大小和質量,低溫條件下在等滲液中破碎細胞,差速離心後,獲得線粒體。經活性染料健那綠Janus green B染色,線粒體呈淺藍色。
三、實驗內容
1.線粒體的分離提取 2. 鼠肝的勻漿製備 3. 線粒體的活體染色
四、實驗步驟
(一)動物組織線粒體的分離,提取與觀察
顯微鏡檢查:將1%Janus green B溶液按1:1比例加入線粒體懸液中,在室溫或水浴中染15~20分鐘,用吸管吸取一滴線粒體懸液,滴於載玻片上,加蓋玻片後,放顯微鏡下進行觀察,線粒體為藍綠色圓形顆粒。
2.組織培養細胞的線粒體的提取與觀察
注意問題
1.整個操作過程為保證線粒體的完整,應儘量使操作時的環境如溫度(0—4℃),pH (7.0左右)保持恆定,同時儘可能短操作時間。
2.組培細胞消化時要特別小心,防止損失或反覆。(損失指細胞脫落到消化液中)。
3.勻漿時,所用的介質一定是等滲緩衝液,常用的有0.25 mol/L蔗糖溶液或生理鹽水代替Hank’s液
4.勻漿次數依照勻漿器的鬆緊而定,次數過少,細胞破損不完全,就會影響線粒體產量。
5.所以取2/3上清夜用來製備線粒體是為防止細胞碎片過多影響觀察。
6.整個分離過程,一般最好在30—60分鐘內完成,不宜過長。
